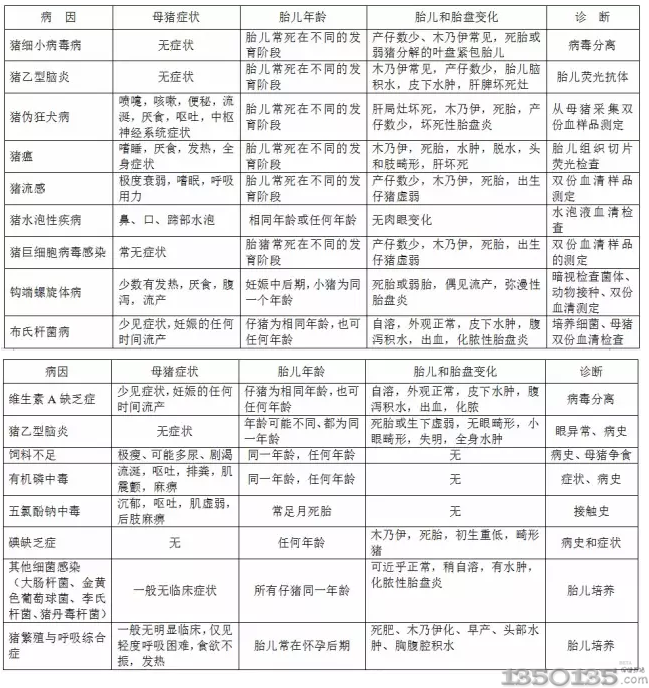
|
 |
3 猪链球菌对大环内酯类耐药的特点
3.1 多重耐药
大环内酯类耐药猪链球菌中多重耐药株(MDR)占很大比例,通常为四耐(同时耐4种药物)以上。药敏试验表明长春地区猪链球菌分离株MLSB类耐药率91%,多重耐药率为(对MLSB类、四环素类、氟喹诺酮类、氯霉素类、氨基糖苷类)82%[4]。王丽平等[19]检测了65株猪源链球菌对32种药物的耐药性,其中大环内酯多重耐药株占70%以上。Martel A等[9]检测了87株猪链球菌(1999年-2000年),71%的菌株对MLSB类耐药,85%的菌株对四环素类耐药。
我国猪链球菌对大环内酯/林可酰胺类耐药性高于丹麦、瑞典、日本等国家,且我国多药耐药情况更为严重[19]。研究表明,细菌长时间承受着多种抗生素选择的压力,因此,只有携带多种耐药决定子的细菌才得以生存,这些耐药决定子一般位于转座子中,可在不同菌种、菌株间传递,造成细菌对抗生素的普遍交叉耐药。Stuart J G等[20]在红霉素、克林霉素、四环素多重耐药猪链球菌中检测到一种类似Tn916的接合型转座子,携带红霉素、四环素等抗性基因。Martel A等[21]在携带ermB基因的猪链球菌中检测到tet基因及intTn1545基因,体外试验发现,ermB、tetO基因一同以较低频率在猪源和人源链球菌之间转移。
3.2 血清型相关性
猪链球菌根据菌体荚膜抗原特性的不同,分成35个血清型。研究发现,大环内酯类耐药性与菌株血清型之间可能存在相关性。
分离自丹麦的猪链球菌对大环内酯类药敏试验发现,1967年-1982年2型分离株全部为敏感,7型23.3%耐药,1992年-1997年2型分离株20.4%耐药,7型44.8%耐药[22]。另一项来自丹麦1995年67例和1996年31例猪链球菌的报道,2型猪链球菌中9.7%对林可霉素耐药,12.9%对螺旋霉素耐药,其他血清型猪链球菌中56.8%对这两种抗菌药物耐药[23]。分离自西班牙的151株猪链球菌,9型猪链球菌对泰乐菌素耐药性显著高于2型猪链球菌(前者耐药率94%,后者耐药率77%)[10]。以上数据,可能反映了菌株间抗生素选择压力及获得耐药性能力的差异。
耐药性与血清型相关性研究对猪链球菌分子流行病学研究具有很重要的意义,此外,耐药性与生化特性、致病性等的关系有待于进一步研究。
4 耐药基因传播机制
分析猪源链球菌耐药性产生的原因,除抗菌药物长期应用导致的“选择性压力”筛选和抗菌药物的不合理使用外,耐药基因在菌株间的广泛传播至关重要。链球菌大环内酯类耐药基因定位于质粒或转座子上,如在A、B、C、D和G群链球菌中发现编码MLSB耐药的质粒(17 Mdal~20 Mdal,同源性78%~95%,接合型或非接合型),宿主范围很宽(可在种内及种间、属间转移)。又如在一株粪链球菌的多重耐药质粒上,MLSB耐药基因定位于一个3.3 Mdal的转座子Tn917上[24]。
细菌耐药可分为天然耐药和获得性耐药。猪链球菌大环内酯类耐药基因erm及mef均为获得性耐药基因,位于转座子内,或由质粒携带,可在细菌间广泛传播。Stuart J G等[20]在大环内酯类耐药猪链球菌中检测到非依赖质粒的接合型转座子(类似Tn916),携带红霉素和四环素抗性并以一定的频率在猪源和人源链球菌间传递。对29株猪扁桃体及鼻腔中不同种属细菌的ermB基因进行测序,其中12株菌的ermB序列相同,其余菌株序列只有1个~6个核苷酸的不同[25]。检测6株猪链球菌ermB基因同源性,与 GenBank中的肺炎链球菌Tn1545转座子、屎肠球菌的质粒pRUM786等的序列同源性为 98%~100%[18]。通过体外试验证明,ermB耐药基因(位于Tn1545上)在猪链球菌和人肺炎链球菌、化脓链球菌等之间以较低的频率进行交换[21]。
畜牧业生产中大量使用抗生素进行动物疾病防治,尤其药物添加剂广泛应用等都促进了细菌耐药的发生与传播。细菌对某种抗生素的固有耐药是其天然特性,耐药变异是细菌适应环境进化的结果。在抗生素选择压力下,质粒和与之相关的转座子使耐药性的转移和传播更加迅速,水平获得耐药基因的亚群可发展成优势菌群并将其垂直遗传给子代。因此,合理使用抗菌药物,加强耐药性监测,开发新药以及疫苗等对于减缓耐药性的产生和传播具有十分重要的意义。